

For the most part, fMRI studies attempt to localize cognitive processes to specific regions in the brain. Popular media often introduce these studies with headlines that tout the discovery of “the brain region” for memory, language, empathy, moral reasoning, loving weiner schnitzel and so on.
These headlines can be terribly misleading, as they’re often misinterpreted to suggest a specific brain region is dedicated to a single function, when, in fact, any given function maps on to a network of regions (forming a circuit), while any given region is part of multiple circuits subserving many functions. Similar faux pas can be found in descriptions of the functions associated with genes, e.g. “The gene for (fill in the blank).”
A few years back, the NY Times ran an infamous piece featuring the work of a neuromarketing company. In a horrible experiment fit for The Onion, participants lay in the scanner while looking at pictures of then presidential candidates. Subjects showed increased amygdala activation to pictures of Mitt Romney, which researchers interpreted as a sign of anxiety.

But after watching Romney speak on video, the amygdala activity died down, which researchers said showed that voters’ anxiety had decreased.
Meanwhile subjects’ anterior cingulates lit up to pictures of Hillary Clinton.

Here’s how researchers interpreted this neural activity:
Emotions about Hillary Clinton are mixed. Voters who rated Mrs. Clinton unfavorably on their questionnaire appeared not entirely comfortable with their assessment. When viewing images of her, these voters exhibited significant activity in the anterior cingulate cortex, an emotional center of the brain that is aroused when a person feels compelled to act in two different ways but must choose one. It looked as if they were battling unacknowledged impulses to like Mrs. Clinton.
The Times article about the “research” was quickly and roundly criticized by prominent neuroscientists, 17 of whom quickly responded with a signed letter to the editor, which the Times ran a couple of days later:
To the Editor:
“This Is Your Brain on Politics” (Op-Ed, Nov. 11) used the results of a brain imaging study to draw conclusions about the current state of the American electorate. The article claimed that it is possible to directly read the minds of potential voters by looking at their brain activity while they viewed presidential candidates.
For example, activity in the amygdala in response to viewing one candidate was argued to reflect “anxiety” about the candidate, whereas activity in other areas was argued to indicate “feeling connected.” While such reasoning appears compelling on its face, it is scientifically unfounded.
As cognitive neuroscientists who use the same brain imaging technology, we know that it is not possible to definitively determine whether a person is anxious or feeling connected simply by looking at activity in a particular brain region. This is so because brain regions are typically engaged by many mental states, and thus a one-to-one mapping between a brain region and a mental state is not possible.As cognitive neuroscientists, we are very excited about the potential use of brain imaging techniques to better understand the psychology of political decisions. But we are distressed by the publication of research in the press that has not undergone peer review, and that uses flawed reasoning to draw unfounded conclusions about topics as important as the presidential election.
Adam Aron, Ph.D., University of California, San Diego
David Badre, Ph.D., Brown University
Matthew Brett, M.D., University of Cambridge
John Cacioppo, Ph.D., University of Chicago
Chris Chambers, Ph.D., University College London
Roshan Cools, Ph.D., Radboud University, Netherlands
Steve Engel, Ph.D., University of Minnesota
Mark D’Esposito, M.D., University of California, Berkeley
Chris Frith, Ph.D., University College London
Eddie Harmon-Jones, Ph.D., Texas A&M University
John Jonides, Ph.D., University of Michigan
Brian Knutson, Ph.D., Stanford University
Liz Phelps, Ph.D., New York University
Russell Poldrack, Ph.D., University of California, Los Angeles
Tor Wager, Ph.D., Columbia University
Anthony Wagner, Ph.D., Stanford University
Piotr Winkielman, Ph.D., University of California, San Diego
Undoubtedly, fewer people saw that letter than saw the original article, which was much more prominently displayed.

(By the above study’s logic, looking at a picture of Donald Trump should elicit activity in the anterior insula, a region often associated with disgust responses)
It’s unfortunate that the study received such a prominent platform for distribution because people, especially non scientists, can be heavily influenced by articles with pictures of brains or technical sounding neuro language. One study, which I’ve written about on the blog, found that people were much more likely to believe a nonsensical article if it had meaningless neuroscience language in it than if it didn’t. As the average lay person doesn’t possess the technical skills to distinguish between valid and invalid fMRI studies, it’s up to the scientific community to police itself, which it does a pretty good job of through the peer review process.
This next study I’ll talk about demonstrates some of the challenges inherent to mapping localized neural activity onto unseen mental processes; in this case, the subjective experience of intense, romantic long-term love.
Aaron and colleagues previously published a study that presented neural correlates of intense romantic love (2005). In brief, the study reported that regions in the reward circuit of the brain were activated in response to pictures of a lover (versus a close friend). In the current study, they wanted to explore if these findings could be extended to long-term married couples (couples together for more than 20+ years who report still being madly in love).
Participants lay in the scanner and were repeatedly presented with pictures from four different categories: their partner, a close friend, and both a highly familiar and low-familiar neutral acquaintance. They were instructed to think about “experiences with each stimulus person (that were) nonsexual in nature.”
The fMRI data was analyzed via subtraction, a common fMRI analysis method in which one condition is compared to another to see if any differences fall out. The contrast of interest was between the partner and the close friend. In a cognitive process sense, the only difference thought to exist between perceiving these two individuals was thought to be the subject’s romantic love for one and not for the other. So if neural activity in the close friend condition is subtracted from activity in the partner condition, whatever is left over should represent the neural substrate of romantic love.
Researchers found activation in the ventral tegmental area, substantia nigra and nucleus accumbens (and the hippocampus, which corresponded with reported sex frequency, but the effect seems to be driven largely by two outliers, one of whom reported they have sex almost every day).
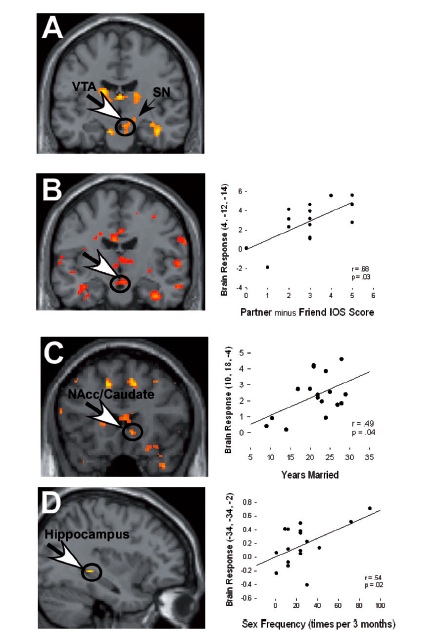
The activity does suggest a classic reward response and replicates previous findings. However, the big question isn’t whether there is a response, but rather what’s driving it?
A valid fMRI study doesn’t only rely on the integrity or analysis of the fMRI data, but, also, and perhaps more importantly, on the experimental design. In order to attribute increased activation in one condition versus the other to a specific cognitive function, one must be confident they have created conditions that have cleanly isolated the independent variable of interest (romantic love). The ventral tegmental area and other regions in the basal ganglia have been repeatedly shown to encode reward value – that is, they respond to things that give us hedonic pleasure, such as food, drugs, sex or receiving money. Past work has shown that intense romantic love is associated with activity in the those regions (Aaron 2005). In the current study, activity in some neural regions previously associated with maternal pair bonding was shown (substantia nigra, for one). The authors hypothesized that neural correlates for romantic long-term love should encompass those associated with both intense romantic love and maternal pair bonding.
But this analysis is dependent on long-term, romantic love being the only difference between conditions that would explain the differences in brain activity. And that may not entirely be the case.
Alternative Explanations
Just to refresh, the major dependent measure of interest was neural activity, especially of reward circuitry, while subjects looked at pictures of their long-term partner versus a close friend. One additional difference between these conditions (beyond romantic love) is that romantic partners are probably more familiar and closer to participants than close friends. This is a shortcoming acknowledged by the authors.
This difference suggests a causal chain of cognitive operations that could offer alternative explanations for some of the data seen in this study. First, It’s been shown that we prefer things that are familiar to us (the “mere exposure” effect, Zajonc, 1968). Second, we’re able to process familiar things (or people) much more fluently compared to the less familiar (Reber). Third, fluency processing has been associated with judgments of aesthetic appreciation such that the more fluently we can process something, the more beautiful or attractive we’re likely to rate it (Alter). Although the objective attractiveness of the photos was controlled for via a group of independent raters, the participants were likely much more subjective in their judgments and perhaps found their partners more attractive than an objective viewer might. Viewing attractive faces has been shown to elicit strong neural activity, particularly in the reward circuitry (NaCC and OFC).
Furthermore, it has been posited that people incorporate close others into their psychological construct of self. Recent studies (deGreck 2008) showed that regions active in a reward task, such as the bilateral ventral striatum, and the ventral tegmental area (VTA), are also involved in differentiating between high and low personal relevance.It seems that we find thinking about ourselves pretty damn rewarding! (We’re all at least a little bit narcissistic). To the extent that someone has been incorporated into our self concept, thinking about that person, or looking at their picture as in this study, could be correlated with responses in reward regions of the brain in part because they activate thoughts of ourselves.
Both the familiarity –> processing fluency –> attractiveness model and self relevant thinking are plausible alternative explanations for at least some of the neural correlates found in this paper.
One other potential area of concern is that there is no way of knowing that participants weren’t thinking about sex with their partners, even though they were told not to. This might be especially difficult to achieve, especially for the two outliers who reported almost daily sex. Regions active during sexual arousal include R. amygdala, hypothalamus, hippocampus, midbrain, mOFC and nucleus accumbens, many of which were found to be active in this study.
The neural activity measured here may very well reflect some aspect of individuals’ love for their partners. But there seem to be other possible explanations for some of the data. I suppose that’s why people call the study of consciousness, of which subjective experiences such as romantic love are a subset, the “hard problem.”
References
Acevedo BP, Aron A, Fisher HE, & Brown LL (2011). Neural correlates of long-term intense romantic love. Social cognitive and affective neuroscience PMID: 21208991
Aron, A., Fisher, H., Mashek, D., Strong, G., Li, H., Brown, L. (2005). Reward, motivation and emotion systems associated with early-stage intense romantic love. Journal of Neurophysiology, 93, 327–37.
DEGRECK, M., ROTTE, M., PAUS, R., MORITZ, D., THIEMANN, R., PROESCH, U., BRUER, U., MOERTH, S., TEMPELMANN, C., & BOGERTS, B. (2008). Is our self based on reward? Self-relatedness recruits neural activity in the reward system NeuroImage, 39 (4), 2066-2075 DOI: 10.1016/j.neuroimage.2007.11.006
Alter, A., & Oppenheimer, D. (2008). Easy on the mind, easy on the wallet: The roles of familiarity and processing fluency in valuation judgments Psychonomic Bulletin & Review, 15 (5), 985-990 DOI: 10.3758/PBR.15.5.985
Peskin, M., & Newell, F. (2004). Familiarity breeds attraction: Effects of exposure on the attractiveness of typical and distinctive faces Perception, 33 (2), 147-157 DOI: 10.1068/p5028
Reber, R., Schwarz, N., & Winkielman, P. (2004). Processing Fluency and Aesthetic Pleasure: Is Beauty in the Perceiver’s Processing Experience? Personality and Social Psychology Review, 8 (4), 364-382 DOI: 10.1207/s15327957pspr0804_3






You must be logged in to post a comment.